

Krystian Florkowski
Zakład Taksonomii Roślin, Wydział Biologii, Uniwersytet im. A. Mickiewicza w Poznaniu
krystian.florkowski@amu.edu.pl
Cytowanie: Florkowski, K. (2018). Wpływ wielkości rozłupek na kiełkowanie i rozwój siewek selerów błotnych Apium repens (Jacq.) Lag. Wiadomości Botaniczne, 62. https://doi.org/10.5586/wb.2018.009

Streszczenie: Dotychczas uznawano, że selery błotne występujące na obszarze Polski na wyspowych, izolowanych od zwartego zasięgu stanowiskach, nie wytwarzają w pełni funkcjonalnych propagul. Tymczasem szczegółowe badania populacyjne przeprowadzone w ostatnich latach w Wielkopolsce dowiodły, że selery błotne wytwarzają dojrzałe nasiona oraz rozmnażają się także generatywnie. W niniejszej pracy przedstawiono wyniki testowania wpływu wielkości nasion i długości ich spoczynku na szybkość kiełkowania oraz tempo wzrostu siewek do etapu wytworzenia pierwszych rozłogów. Eksperyment prowadzony był równolegle w kontrolowanych warunkach środowiskowych w komorze hodowlanej oraz w warunkach polowych (inspekt otwarty) na terenie Ogrodu Botanicznego Uniwersytetu im. Adama Mickiewicza w Poznaniu. W eksperymencie wykorzystano rozłupki pochodzące z trzech największych populacji Wielkopolski (Brenno 2, Brenno Ostrowo i Ostrowo Hutka) zebrane w warunkach in situ oraz hodowli w ogrodzie botanicznym. Stwierdzono, że wielkość nasion ma wpływ na efektywność kiełkowania oraz rozwój siewek selerów błotnych Apium repens. Przeprowadzona korelacja r-Pearsona dowiodła, że wielkość rozłupek ma pozytywny wpływ na szybkość kiełkowania oraz tempo wzrostu siewek. Czas wymagany do kiełkowania rozłupek przechowywanych przez okres jednego roku od zebrania sięgał od blisko 30 dni dla próby w fitotronie do około 190 dni w warunkach polowych. Uzyskana wiedza z zakresu biologii rozwoju ma wartość aplikacyjną w aktywnej ochronie selerów błotnych.
The effect of seed size on germination and growth of seedlings of creeping marshwort Apium repens (Jacq.) Lag
Abstract: According to previous studies it was believed that creeping marshwort occurring in Poland on isolated locations, satellite to the main range of the species, does not produce fully functional propagules. Meanwhile, detailed population studies conducted in recent years in Wielkopolska region have shown that Apium repens produces mature seeds and reproduces generatively. The paper presents the results of testing the effect of seed size and length of their dormancy on the germination rate and the rate of growth of seedlings to the stage with first stolons. The experiment was conducted under controlled environmental conditions: in a breeding chamber (phytotron) and in the field (open hotbed) at the Botanical Garden of the University of Adam Mickiewicz in Poznań. In experiment schizocarps from the three largest populations of Wielkopolska were used (Brenno 2, Brenno Ostrowo and Ostrowo Hutka), collected in situ and cultivated in the botanical garden. Seed size influenced efficiency of germination and seedling development of Apium repens. The r-Pearson correlation proved that fruit size has a positive effect on the germination rate and growth rate of seedlings. The time required for germination of seeds stored for one year after harvesting ranged from nearly 30 days for the phytotron test to about 190 days in field conditions. Acquired knowledge in developmental biology of creeping marshwort has an application value for active conservation of the species.
Keywords: Apium repens; seed size; germination; growth; seedlings
Wprowadzenie
Energia pozyskiwana w procesie fotosyntezy podlega relokacji w różnych organach w zależności od zapotrzebowania na zasilenie podstawowych procesów życiowych, np. wzrostu i rozwoju roślin (Bloom i in., 1985; Hutchings, 2004; Lohier i in., 2014). W reakcji na zmianę warunków środowiska rośliny mogą zmieniać wzorzec alokacji zasobów, w postaci strategii allometrycznej swoich części. Lokacja zasobów jest zależna od stadium ontogenezy i fenotypu będącego odpowiedzią rośliny na warunki środowiskowe oraz konkurencję (Muller i in., 2000; Niu i in., 2008). W zależności od warunków środowiskowych relokacja zasobów energetycznych obejmuje też owoce i nasiona (Mc Connaughat, Coleman, 1999; Porter, Nagel, 2000; Hermans i in., 2006). Uważa się, że zdolność kiełkowania nasion zależy głównie od stanu ich dojrzałości, natomiast o rozwoju siewek decyduje ilość zgromadzonych zasobów energetycznych w nasionach (Eriksson, 1999; Lehtila, Ehrlen, 2005; Souza, Fagundes, 2014; Xiao i in., 2015).
Dotychczas uważano, że selery błotne Apium repens rozmnażają się głównie wegetatywnie za pomocą rozłogów zakorzeniających się w węzłach. Stwierdzano, że pomimo intensywnego kwitnienia, nie zawiązują owoców lub też owoce nie osiągają pełnej dojrzałości na obszarze Polski (Żukowski i in., 1988; Chmiel i in., 2014). Ostatnie obserwacje poczynione w warunkach in situ oraz w trakcie hodowli A. repens w Ogrodzie Botanicznym Uniwersytetu im. Adama Mickiewicza w Poznaniu dowiodły, że w warunkach klimatycznych Polski mogą się formować w pełni dojrzałe rozłupki (Florkowski, 2017; Chmiel, 2016).
W warunkach eksperymentalnych istnieje możliwość poznania wpływu czynników środowiskowych na proces kiełkowania nasion oraz wzrost i rozwój siewek roślin (Useche, Shipley, 2010a, 2010b; Poorter i in., 2012). Celem zaplanowanej serii analiz było wyjaśnienie następujących problemów:
- czy i w jakim stopniu wielkość owoców (nasion) ma wpływ na zdolność kiełkowania oraz wzrost siewek do momentu wytworzenia rozłogów?
- czy sygnałem do wytwarzania rozłogów jest osiągnięcie określonej biomasy roślin?
Przed wyjaśnieniem powyższych problemów badawczych postawiono następujące hipotezy:
- początek wytwarzania rozłogów przez repens determinowany jest osiągnięciem przez siewkę określonej masy,
- wielkość nasion pozytywnie wpływa na szybkość kiełkowania i wzrost siewek,
- owoce (nasiona) repens pozyskane w warunkach in situ z terenu Wielkopolski i w warunkach ex situ na terenie Ogrodu Botanicznego cechują się podobną żywotnością.
Materiał i metody badań
Zadanie badawcze realizowano w kilku etapach obejmujących: zbiór owoców, ich pomiary, przygotowanie podłoża, kiełkowanie, obserwacje wzrostu i rozwoju oraz pomiar suchej masy roślin selerów błotnych (Ryc. 1).
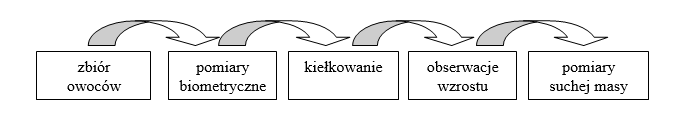
Ryc. 1. Schemat procedury badań.
W eksperymencie wykorzystano rozłupki A. repens zebrane w roku 2015 na obszarze Wielkopolski z populacji: Brenno 2 (B2), Brenno Ostrowo (BO) i Ostrowo Hutka (OH). Losowy pobór rozłupek z różnych części populacji zapewnił uzyskanie próby reprezentatywnej. W tym samym roku pozyskano też owoce z kolekcji ex situ założonych na bazie materiału roślinnego (ramet) pobranych z tych samych stanowisk. Rozłupki były przechowywane w papierowych kopertach w temperaturze pokojowej (20–22°C) w suchym pomieszczeniu. Przy użyciu mikroskopu stereoskopowego wyposażonego w program Cell^A wykonano pomiary długości oraz szerokości wszystkich rozłupek wykorzystanych w eksperymencie. Dla zapewnienia stosowalności jednoczynnikowej analizy wariancji ANOVA rozłupki były pobierane poprzez losowanie niezależne. Analizę korelacji r-Pearsona zastosowano w określeniu wpływu wielkości rozłupek na masę podziemnych i nadziemnych części rośliny.
Rozłupki selerów błotnych poddano kiełkowaniu w kwietniu 2016 roku w stałych warunkach w komorze fitotronowej w trzech powtórzeniach po 30 rozłupek z każdej populacji (Tab. 1). W tej części eksperymentu badany był wpływ wielkości rozłupek, powstałych w warunkach in situ i ex situ, na szybkość i siłę kiełkowania oraz tempo rozwoju siewki do fazy pierwszego liścia. Młode siewki w fazie wytworzenia pierwszych liści zostały przesadzone do donic i przeniesione do otwartego inspektu w ogrodzie botanicznym.
Tab. 1. Parametry fizyczne warunków kiełkowania nasion w fitotronie.
| Pora dnia / godzina | Dzień 4.00–20.00 | Noc 20.00–4.00 |
| Oświetlenie | 600 µMol/m2s | 0 µMol/m2s |
| Temperatura | 21°C | 15°C |
| Wilgotność | 60% | 68% |
W listopadzie 2016 roku rozpoczęto kolejną część eksperymentu, obejmującą porównanie wpływu warunków polowych i panujących w fitotronie na zdolność kiełkowania, wzrost do fazy pierwszego liścia oraz czas potrzebny do wytworzenia rozłogów z określeniem wielkości suchej masy uzyskanej przez rośliny w tej fazie rozwoju. W tym przypadku w testowaniu zdolności kiełkowania nasion wykorzystano łącznie 160 sztuk rozłupek pobranych w terenie z dwóch stanowisk: Brenno Ostrowo i Ostrowo Hutka, na których A. repens tworzy największe w Wielkopolsce populacje (Rogozińska, 2012). Każda rozłupka użyta do eksperymentu została zmierzona pod względem długości oraz szerokości. Rozłupki wysiano w dwóch równoległych eksperymentach: w komorze fitotronowej oraz w otwartym inspekcie po 40 sztuk z każdego stanowiska. Rozłupki przeznaczone do prób w komorze fitotronowej (wysiane na początku kwietnia 2017 roku) poddane były wcześniej stratyfikacji w temperaturze 3°C przez okres 3 dni. Rozłupki wysiane w listopadzie 2016 roku w otwartym inspekcie poddane były niskiej temperaturze w okresie zimowym. Wysiew w inspekcie i w fitotronie przeprowadzono w identycznym podłożu: ziemi parowanej o ustalonej wcześniej zawartości głównych składników mineralnych (Tab. 2).
Tab. 2. Parametry chemiczne podłoża.
| Wyszczególnienie parametrów chemicznych | Wartość parametru |
|---|---|
| pH w H2O | 5,0–6,5 |
| Azot w mg/l | 150–250 |
| Fosfor w mg/l | 100–200 |
| Potas w mg/l | 250–350 |
| Zasolenie w g KCl | 0,8–1,5 |
| Mikroelementy | Mg, Fe, Mn, Zn, B, S |
Obserwacje wzrostu i rozwoju, zarówno w inspekcie, jak i w komorze fitotronowej trwały od dnia wysiewu do wytworzenia pierwszych rozłogów z kątów liściowych. Siewki na etapie inicjacji wytwarzania pędów bocznych zostały pobrane z kuwet i oczyszczone z ziemi. Rozdzielone części podziemne i nadziemne rośliny wysuszono w temperaturze pokojowej, oraz zważono na wadze laboratoryjnej. Zapis kiełkowania oraz wzrostu siewek prowadzony był w odstępie 48 godzin.
W czasie eksperymentu zwracano uwagę na osiąganie poszczególnych faz rozwojowych:
- Faza I – od pęknięcia łupiny nasiennej do początku wzrostu korzenia zarodkowego
- Faza II – wzrost siewki do momentu zrzucenia łupiny nasiennej i rozpostarcia liścieni
- Faza III – wzrost z wykształconymi liścieniami
- Faza IV – wytworzenia pierwszego liścia.
Kolejne fazy rozwoju liczone były jako pojawianie się kolejnych liści.
Wyniki
Przeprowadzona analiza statystyczna przy pomocy testu jednoczynnikowej ANOVA wykazała istotne statystycznie różnice, odnoszące się do długości owoców pochodzących z poszczególnych populacji ex situ F(2, 87) = 28,96; p < 0,001; η2 = 0,40 oraz szerokości F(2, 87) = 20,98; p < 0,001; η2 = 0,33. Jednoczynnikowa analiza wariancji dla rozłupek pochodzących z populacji in situ również wykazała istotne statystycznie różnice dotyczące długości F(2, 87) = 42,15; p < 0,001; η2 = 0,49 oraz szerokości F(2, 87) = 5,49; p < 0,001; η2 = 0,112. Największy obserwowany efekt zróżnicowania dotyczy długości rozłupek w populacji in situ – wyjaśnia aż 49% zmienności. Tymczasem parametr średniej szerokości rozłupek in situ wyjaśnia zaledwie 11,2% całej zmienności między populacjami.
Przeprowadzony test post hoc wskazuje, że wszystkie rozłupki w serii zarówno in situ, jak i ex situ różnią się między sobą długością. Najdłuższe rozłupki odnotowano w populacji Brenno Ostrowo in situ (MS = 1108,04; SD = 82,71), a największa różnica w długości rozłupek, istotna statystycznie, miała miejsce w odniesieniu do populacji z Ostrowo Hutki (MS = 929,39; SD = 60,16), p < 0,001; d Cohena = 2,51; 95%CIróżnicy[115,11; 242,91].
Dla tych samych populacji różnice w średnich wartościach szerokości nie okazały się istotne statystycznie (p > 0,05). Największa szerokość rozłupek odnotowana została w puli z populacji Brenno Ostrowo ex situ (Tab. 3). Tylko rozłupki z populacji Ostrowo Hutka wykazują istotną różnicę statystyczną względem serii z innych populacji. W serii in situ rozłupki z Ostrowa Hutki (MS = 624,47; SD = 66,95) różnią się najsilniej względem populacji z Brenna 2 (MS = 688,17; SD = 58,24), p < 0,01; d Cohena = 1,02; 95%CIróżnicy[8,84; 118,55], natomiast w serii ex situ rozłupki wywodzące się z Ostrowa Hutki (MS = 604,39; SD = 90,93) różnią się najsilniej względem rozłupek z populacji Brenno Ostrowo (MS = 707,13; SD = 52,73), p < 0,001; d Cohena = 1,47; 95%CIróżnicy[47,89; 157,5]. Porównując owoce z populacji in situ i ex situ nie stwierdzono statystycznie istotnego wpływu warunków środowiskowych na wielkość rozłupek.
Stwierdzone różnice w zdolności kiełkowania nie są skorelowane z wielkością rozłupek. Najwięcej skiełkowanych nasion odnotowano w serii pochodzących z populacji ex situ Brenno Ostrowo (BO) oraz Ostrowo Hutka (OH). W przypadku owoców zebranych z populacji in situ jedynie rozłupki pochodzące z populacji Brenno 2 (B2) wykazały większy odsetek skiełkowania (Tab. 3). Średni czas potrzebny do wykiełkowania wynosił 8,6 dnia. Najszybciej kiełkowały rozłupki pochodzące ze stanowisk ex situ BO (7,6 dnia) i B2 (7,9 dnia), a najwolniej z ex situ OH (10,1 dnia) oraz in situ B2 (10,8 dnia). Statystycznie istotnie wolniej kiełkowały rozłupki w populacji in situ B2 (MS = 10,80; SD = 2,78) względem populacji ex situ BO (MS = 7,57; SD = 1,04); p < 0,05; d Cohena = 1,69; 95%CIróżnicy[2,11; 4,35] oraz ex situ B2 (MS = 7,91; SD = 1,90); p < 0,05; d Cohena = 1,23; 95%CIróżnicy[1,43; 4,34].
Podczas obserwacji wzrostu zanotowano różnice w tempie wytworzenia pierwszych liści (faza 4) między populacjami ex situ a in situ. Największa różnica istotna statystycznie wyniosła dla populacji B2 (MS = 23,6; SD = 1,75); (MS = 12,70; SD = 3,20); p < 0,001; d Cohena = 4,39; 95%CIróżnicy[8,45; 13,28] oraz OH (MS = 19,88; SD = 1,03), (MS = 11,87; SD = 4,16); p < 0,001; d Cohena = 3,09; 95%CIróżnicy[5,76; 10,27].
Tab. 3. Wyniki pomiarów wielkości owoców pobranych w warunkach in situ i ex situ i wybranych parametrów kiełkowania i rozwoju siewek w warunkach fitotronu.
| Pochodzenie owoców | Wyszczególnienie analizowanych cech | Wartość cech morfometrycznych owoców i efektu kiełkowania | ||
|---|---|---|---|---|
| Brenno Ostrowo | Brenno 2 | Ostrowo Hutka | ||
| Ex situ | % wykiełkowanych nasion | 100% | 70% | 87% |
| dni kiełkowania | 7,57 ±1,04 | 7,91 ±1,90 | 9,96 ±3,10 | |
| dni w fazie
pierwszego liścia |
19,17 ±1,34 | 23,6 ±1,75 | 19,88 ±1,03 | |
| µm ±SD długość | 1103,31 ±104,31 | 1008,3 ±71,15 | 926,86 ±90,63 | |
| µm ±SD szerokość | 707,13 ±52,73 | 698,35 ±53,49 | 604,39 ±90,93 | |
| In situ | % wykiełkowanych nasion | 63,3% | 85% | 77% |
| dni kiełkowania | 8,79 ±1,32 | 10,8 ±2,78 | 8,13 ±1,74 | |
| dni w fazie
pierwszego liścia |
14,74 ±2,02 | 12,7 ±3,2 | 11,87 ±4,16 | |
| µm ±SD długość | 1108,4 ±82,71 | 1032,98 ±82,39 | 929,39 ±60,16 | |
| µm ±SD szerokość | 658,23 ±93,72 | 688,17 ±58,24 | 624,47 ±66,95 | |
W kolejnym eksperymencie testowano wpływ warunków polowych i fitotronowych na efektywność kiełkowania rozłupek. Okazało się, że rozłupki umieszczone w komorze fitotronowej kiełkowały zdecydowanie lepiej w porównaniu do rozłupek posianych w otwartym inspekcie (Tab. 4). Owoce z populacji użytych w eksperymencie, tj. Brenno Ostrowo i Ostrowo Hutka różniły się statystycznie pod względem długości na poziomie F(1, 158) = 178,96; p < 0,001; η2 = 0,53 i szerokości F(1, 158) = 264,48; p < 0,001; η2 = 0,63. Odsetek kiełkujących rozłupek był bardzo zbliżony do efektywności kiełkowania w pierwszym eksperymencie. Zdecydowanie niższą efektywność kiełkowania uzyskano w warunkach otwartego inspektu.
Po rocznym przechowywaniu owoców w temperaturze pokojowej (20–22°C) przy braku dostępu światła, średnia liczba dni potrzebnych do skiełkowania nasion w komorze fitotronowej pochodzących ze stanowiska Brenno Ostrowo była większa prawie o 20 dni (MS = 27,91; SD = 17,18 dni) od poprzedniej próby (MS = 8,13; SD = 1,74 dni), natomiast w próbie pochodzącej ze stanowiska Ostrowo Hutka o 30 dni (MS = 38,96; SD = 37,28 dni), (MS = 8,79; SD = 1,32 dni). Dodatkowo przedział czasowy kiełkowania siewek wyniósł od 3 do 115 dni dla OH oraz od 13 do 104 dni dla BO od wysiewu, co finalnie przełożyło się na wyższe odchylenie standardowe. Dla porównania w pierwszym eksperymencie czas kiełkowania oscylował wokół 10 dni od wysiewu a największa różnica w tempie kiełkowania wynosiła od 7 do 21 dni i dotyczyła populacji ex situ OH. Kiełkowanie rozłupek w otwartym inspekcie przypadało na połowę maja (około 6,5 miesiąca od wysiewu), gdy warunki pogodowe sprzyjały rozwojowi siewek.
Znając długość i szerokość każdej rozłupki podjęto próbę skorelowania tych cech z niektórymi elementami wczesnego wzrostu i rozwoju selerów błotnych. Bez względu na pochodzenie owoców, szybkość kiełkowania liczona w dniach dla każdej rozłupki była pozytywnie skorelowana z długością oraz szerokością rozłupek, tzn. wraz z wielkością rozłupek wzrasta liczba dni potrzebnych do skiełkowania. Zaistniałą sytuację obrazuje wskaźnik r-Pearsona, który wynosi odpowiednio r = 0,338 dla p = 0,010 (długość) i r = 0,343 dla p = 0,009 (szerokość), wskazując na umiarkowany stopień korelacji. Można by przypuszczać, że siewki powstałe z dużych rozłupek, pomimo dłuższego okresu spoczynku, będą szybciej wzrastać dzięki większej ilości zmagazynowanych asymilatów, czyli cechować się ujemną korelacją pomiędzy fazą wzrostu a liczbą dni, które upłynęły od wykiełkowania. Podczas eksperymentu rezultat był odwrotny i dowodził silnej pozytywnej korelacji czasu osiągania kolejnych faz rozwojowych do liczby dni potrzebnych do określonej fazy (r = 0,723 dla p < 0,000).
Ponadto nie stwierdzono zależności wielkości (suchej) masy uzyskanej w fazie wytworzenia zawiązku pędów bocznych od wielkości rozłupek (r = 0,0918 dla p = 0,511). Taki wynik może świadczyć, że siewki powstające z małych nasion osiągają porównywalną masę do siewek powstałych z większych nasion w chwili zawiązania pędu bocznego. Początek tworzenia się pędów bocznych następuje w przedziale 0,04-0,1 mg wielkości suchej masy uzyskanej przez rośliny.
Dodatkowo dokonano równolegle pomiaru masy części podziemnej oraz nadziemnej. Okazało się, że selery błotne wytwarzają bogatą sieć korzeni o wiązkowym charakterze, których biomasa względem części nadziemnej wyraża się w proporcji 1:2, wykazując bardzo silną korelację względem siebie (r = 0,709 dla p < 0,00).
Tab. 4. Porównanie efektów kiełkowania i rozwoju siewek w warunkach fitotronu i w warunkach polowych.
| Warunki kiełkowania | Wyszczególnienie analizowanych cech | Wartość cech morfometrycznych owoców i efektu kiełkowania | |
|---|---|---|---|
| Ostrowo Hutka | Brenno Ostrowo | ||
| Polowe (otwarty inspekt) | % wykiełkowanych nasion | 15% | 10% |
| x̄ dni kiełkowania | 195,33 ±14,46 | 192,67 ±12,36 | |
| x̄ dni w fazie pierwszego liścia | 15,28 ±10,28 | 13,11 ±9,38 | |
| x̄ µm ±SD długość | 978,442 ±48,549 | 1075,61 ±43,699 | |
| x̄ µm ±SD szerokość | 680,608 ±43,130 | 784,061 ±35,294 | |
| Komora fitotronowa | % wykiełkowanych nasion | 80% | 62,5% |
| x̄ dni kiełkowania | 38,96 ±37,28 | 27,91 ±17,18 | |
| x̄ dni w fazie pierwszego liścia | 10,12 ±3,11 | 9,91 ±2,19 | |
| x̄ µm ±SD długość | 961,233 ±64,19 | 1096,934 ±59,197 | |
| x̄ µm ±SD szerokość | 680,905 ±51,085 | 798,593 ±40,829 | |
Dyskusja
Wyniki przeprowadzonego eksperymentu, dotyczącego kiełkowania oraz najwcześniejszych faz rozwoju Apium repens, potwierdziły znane zależności szybkości wzrostu siewek od środowiska w jakich doszło do skiełkowania. Warunki środowiskowe: temperatura, wilgotność oraz skład mineralny gleby, to istotne czynniki wpływające na kiełkowanie roślin (Milberg, Lamont, 1997; Muller i in., 2000; Gutterman, 2000; Souza, Fagundes, 2014). Badania przeprowadzone przez Burmeier i Jensen (2008) udowodniły, że ważnym czynnikiem kiełkowania Apium repens jest światło. Drastyczne różnice w liczebności skiełkowanych nasion w komorze fitotronowej względem kiełkujących w warunkach polowych mogą wynikać ze zmienności czynników środowiskowych, zwłaszcza atmosferycznych. Stałe warunki w komorze fitotronowej sprzyjają rozwojowi siewek podczas kiełkowania. W środowisku naturalnym oraz w inspekcie polowym nasiona oraz delikatne siewki narażone są na zmienne warunki atmosferyczne, jak np. gwałtowny deszcz czy obniżona temperatura, mogące spowodować obumarcie młodych roślin lub zarodka, który nie zdążył wytworzyć korzenia zarodkowego (Debeaujon, 2000).
W pracach eksperymentalnych często podejmowanym zagadnieniem jest testowanie wpływu długości okresu spoczynkowego nasion na zdolność ich kiełkowania (Gutterman, 2000; Ouled Belgacem i in., 2006). Nasiona wielu gatunków roślin w odpowiednich warunkach przetrwać mogą wiele lat, co pokazały badania prowadzone na przełomie XIX i XX wieku (Darlington, Steinbauer, 1961; Telewski, Zeevaart, 2002). Podczas prac terenowych zaobserwowano, nierzadko w ciągu jednego roku występowanie młodych siewek Apium repens w aluwiach, odsłoniętych brzegach jezior oraz miejscach, gdzie uważano je za wymarłe (Chmiel, 2016). Obserwacje te mogą świadczyć o tym, że nasiona selerów błotnych mogą czekać na odpowiednie dla siebie warunki do wzrostu. Dowodem długiej żywotności nasion Apium repens może być informacja z Francji o kiełkowaniu rozłupek po 75 latach (Munoz, 2003).
Podczas eksperymentu zapewnienie jednakowych warunków kiełkowania umożliwiło określenie wpływu wielkości i czasu spoczynku nasion na zdolność kiełkowania oraz przebieg wczesnych etapów rozwoju. Tego rodzaju analizy nie były dotąd brane pod uwagę w pracach eksperymentalnych dotyczących biologii rozwoju A. repens. Udało się opisać zależność szybkości kiełkowania i rozwoju siewek od wielkości rozłupek, lecz wynik był odwrotny do sformułowanej hipotezy, co doskonale obrazuje przeprowadzona korelacja ogólna. Siewki selerów błotnych szybciej kiełkują i rozwijają się z mniejszych nasion, potwierdzając podobne zależności podawane dla innych gatunków (Murali, 1997; Souza, Fagundes, 2014), choć takie zależności nie są regułą wśród roślin (Grzesiuk, 1967; Eriksson, 1999; Lehtila, Ehrlen, 2005; Akinyosoye, 2014). Nasiona mniejsze mają cieńsze łupiny nasienne lepiej przepuszczające wodę, co w konsekwencji przekłada się na szybsze ich pęcznienie (Beninger i in., 1998; Souza, 2001). Zróżnicowanie w tempie kiełkowania może mieć pozytywne znaczenie ekologiczne. Nasiona mniejsze mogą sprzyjać szybkiej rekolonizacji mikrosiedlisk inicjalnych pojawiających się losowo i przejściowo, np. odsłaniających się w okresie wiosennym, wraz z obniżaniem się poziomu lustra wody w jeziorze, aluwiach nadbrzeżnych. Z kolei siewki rozwijające się z dużych nasion, dzięki zgromadzonemu materiałowi, są lepiej przystosowane do konkurowania z innymi gatunkami roślin w zwartych płatach roślinnych (Dolan, 1984; Cordazzo, 2002; Souza, Fagundes, 2014).
Podsumowanie i wnioski końcowe
Rozłupki Apium repens niezależnie od powstania w warunkach ex situ lub in situ cechują się dużą żywotnością. Analizy pokazały, że warunki zewnętrzne mają kluczowy wpływ na zdolność kiełkowania oraz rozwój siewek. W części eksperymentu polegającej na wyeliminowaniu różnic w oddziaływaniu czynników zewnętrznych można było zauważyć, że zróżnicowanie wielkości rozłupek, a przede wszystkim ich długości, wpływa na tempo rozwoju siewek. Szybkość kiełkowania i tempo wzrostu siewek mają kluczowe znaczenie w konkurowaniu z innymi gatunkami o mikrosiedliska powstające niekiedy okazjonalnie w naturalnym środowisku. Selery błotne dobrze radzą sobie z częstym zalewaniem wodą, wydeptywaniem, czy też podkaszaniem i zgryzaniem przez zwierzęta (Burmeier, Jensen, 2009). Szybki wzrost siewek i wczesne wytwarzanie rozłogów skutkuje tym, że selery szybciej kolonizują mikrosiedliska.
Interesującym elementem strategii Apium repens jest rozciągnięcie w czasie kiełkowania nasion po dłuższym ich spoczynku, co w perspektywie zmieniających się warunków może być wyrazem strategii dostosowania kiełkowania do warunków im sprzyjających (wyczekiwania na pojawienie się mikrosiedlisk) i korzystnie wpływających na dalszy rozwój siewek. Wiedza z zakresu biologii rozwoju selerów błotnych we wczesnych fazach jest niezbędna w trafnym doborze metod odtwarzania, czy też zasilania populacji poprzez wysiew, bez konieczności prowadzenia hodowli namnożeniowej.
Bibliografia
Akinyosoye, S. T., Adetumbi, J. A., Amusa, O. D., Olowolafe, M. O. & Olasoji, J. O. (2014). Effect of seed size on in vitro seed germination, seedling growth, embryogenic callus induction and plantlet regeneration from embryo of maize (Zea mays L.) seed. Nigerian Journal of Genetics, 28(2), 1–7. doi: 10.1016/j.nigjg.2015.06.001
Bloom, A. J., Chapin, S. F. & Mooney, A. H. (1985). Resource limitation in plants – an economic analogy. Annual Review of Ecology and Systematics, 16, 363–392. doi: 10.1146/annurev.ecolsys.16.1.363
Burmeier, S. & Jensen, K. (2009). Experimental ecology and habitat specificity of the endangered plant Apium repens (Jacq.) Lag. at the northern edge of its range. Journal Plant Ecology & Diversity, 2, 65–75. doi: 10.1080/17550870902773658
Burmeier, S. & Jensen, K. (2008). Is the endangered Apium repens (Jacq.) Lag. rare because of a narrow regeneration niche?. Plant Species Biology, 23, 111–118. doi: 10.1111/j.1442-1984.2008.00212.x
Chmiel, J., Jackowiak, B. & Ziarnek, K. (2014). Apium repens (Jacq.) Lag. – selery błotne (pęczyna błotna). W: R. Kaźmierczakowa, K. Zarzycki & Z. Mirek (eds). W: Polish red data book of plants. Pteridophytes and flowering plants. (s. 360–362). Kraków: Instytut Ochrony Przyrody Polskiej Akademii Nauk. ISBN 978-83-61191-88-9
Chmiel, J. (2016). Regionalna strategia zarządzania zasobami selerów błotnych Apium repens (Jacq.) Laq. w Wielkopolsce. Dokumentacja wykonawcza (Julian Chmiel). Poznań: Zakład Taksonomii Roślin. Wydział Biologii UAM.
Cordazzo, C. V. (2002). Effect of seed mass on germination and growth in three dominant species in Southern Brazilian coastal dunes. Brazilian Journal of Biology, 62, 427–435. doi: 10.1590/S1519-69842002000300005
Darlington, H. T. & Steinbauer, G. P. (1961). The eighty-year period for dr. Beal’s seed viability experiment. American Journal of Botany, 48(4), 321–325. doi: 10.1002/j.1537-2197.1961.tb11645.x
Debeaujon, I., Léon-Kloosterziel, K. M. & Koornneef, M. (2000). Influence of the testa on seed dormancy, germination, and longevity in Arabidopsis. Plant Physiology, 122, 403–414. doi: 10.1104/pp.122.2.403
Dolan, R. W. (1984). The effect of seed size and maternal source on individual size in a population of Ludwigia leptocarpa (Onagraceae). American Journal of Botany, 71, 1302–1307. doi: 10.2307/2443655
Eriksson, O. (1999). Seed size variation and its effect on germination and seedling performance in the clonal herb Convallaria majalis. Acta Oecologica, 20, 61–66. doi: 10.1016/S1146-609X(99)80016-2
Florkowski K. (2017). Morphological variability of fruits in Apium repens (Jacq.) Lag. – a critically endangered species in Poland. Steciana, 21(3), 85–92.
doi:10.12657/steciana.021.009
Grzesiuk, S. (1967). Fizjologia nasion. Warszawa: Państwowe Wydawnictwo Rolnicze i Leśne.
Gutterman, Y. (2000). Maternal effects on seeds during development. W: M. Fenner (red.), Seeds: The Ecology of Regeneration in Plant Communities, 2nd edition (s. 59-84). United Kingdom: CABI Publishing. doi: 10.1079/9780851994321.0000
Hermans, C., Hammond, J. P., White, P. J. & Verbruggen, N. (2006). How do plants respond to nutrient shortage by biomass allocation? Trends Plant Science, 11(12),610–617.
doi: 10.1016/j.tplants.2006.10.007
Hutchings, M. J. & John, E. A. (2004). The Effects of Environmental Heterogeneity on Root Growth and Root/Shoot Partitioning. Annals of Botany, 94, 1–8.
doi:10.1093/aob/mch111
Lehtila, A. K. & Ehrel, J. (2005). Seed size as an indicator of seeds quality: a case study of Primula veris. Acta Oecologica, 28(3), 207–212. doi: 10.1016/j.actao.2005.04.004
Lohier, T., Jabot, F., Meziane, D., Shipley, B., Reich, P. B. & Deffuant, G. (2014). Explaining ontogenetic shifts in root-shoot scaling with transient dynamics. Annals of Botany, 114(3), 513–524. doi: 10.1093/aob/mcu128
Mc Connaughay, K. D. M. & Coleman, J. S. (1998). Can plants track changes in nutrient availability via changes in biomass partitioning? Plant and Soil, 202(2), 201–209.
doi: 10.1023/A:1004341731703
Milberg, P., & Lamont, B. B. (1997). Seed/cotyledon size and nutrient content play a major role in early performance of species on nutrient-poor soils. New Phytologist, 137, 665–672. doi: 10.1046/j.1469-8137.1997.00870.x
Muller, I., Schmid, B. & Weiner, J. (2000). The effect of nutrient availability on biomass allocation patterns in 27 species of herbaceous plants. Perspectives in Plant Ecology, Evolution and Systematics, 3(2), 115–127.doi: 10.1078/1433-8319-00007
Munoz, F. (2003). Réapparition d’une station de Apium repens (Jacq.) Lagasca dans le Bas-Dauphiné (Isère). Le Monde des Plantes, 481, 11–13. Pozyskano z https://www.researchgate.net/publication/253340210
Murali, K. S. (1997). Patterns of seed size, germination and seed viability of tropical tree species in Southern India. Biotropica, 29, 271–279. doi: 10.1111/j.1744-7429.1997.tb00428.x
Niu, K., Luob, Y., Cholerc, Ph., Dua, G. (2008). The role of biomass allocation strategy in diversity loss due to fertilization. Basic and Applied Ecology, 9, 485–493.
doi: 10.1016/j.baae.2007.06.015
Ouled Belgacem, A., Neffati, M., Papanastasis, V. P. & Chaieb, M. (2006). Effects of seed age and seeding depth on growth of Stipa lagascae R. & Sch. seedlings. Journal of Arid Environments, 65(4), 682–687. doi: 10.1016/j.jaridenv.2005.10.001
Poorter, H. A. & Nagel, O. (2000). The role of biomass allocation in the growth response of plants to different levels of light, CO2, nutrients and water: a quantitative review. Australian Journal of Plant Physiology, 27, 595–607. doi: 10.1071/PP99173
Poorter, H. A., Niklas, K. J., Reich, P. B., Oleksyn, J., Poot, P. & Mommer, L. (2012). Biomass allocation to leaves, stems and roots: meta-analyses of interspecific variation and environmental control. New Phytologist, 193(1): 30–50. doi: 10.1111/j.1469-8137.2011.03952.x
Rogozińska A. (2012). Selery błotne (Apium repens) w Przemęckim Parku Krajobrazowym. W: J. Karg (red.), Biuletyn Parków Krajobrazowych Wielkopolski, 18(20), (s. 29–48). Poznań. ISSN 1426-7756.
Souza, M. L. & Fagundes, M. (2014). Seed size as key factor in germination and seedling development of Copaifera langsdorffii (Fabaceae). American Journal of Plant Sciences, 5, 2566–2573. doi: 10.4236/ajps.2014.517270
Telewski, F. W. & Zeevaart, J. A. (2002). The 120-yr period for Dr. Beal’s seed viability experiment. American Journal of Botany, 89(8), 1285–1288. doi:10.3732/ajb.89.8.1285
Useche, A. & Shipley, B. (2010). Plasticity in relative growth rate after a reduction in nitrogen availability is related to root morphological and physiological responses. Annals of Botany, 106(4), 617–625. doi:10.1093/aob/mcq144
Useche, A. & Shipley, B. (2010). Interspecific correlates of plasticity in relative growth rate following a decrease in nitrogen availability. Annals of Botany, 105(2), 333–339.
doi: 10.1093/aob/mcp284
Xiao, Z., Zhang, Z. & Krebs, Ch. J. (2015). Seed size and number make contrasting predictions on seed survival and dispersal dynamics: A case study from oil tea Camellia oleifera. Forest Ecology and Management, 343, 1–8. doi: 10.1016/j.foreco.2015.01.019
Żukowski, W., Latowski, K., Jackowiak, B. & Chmiel, J. (1988). Apium repens (Jacq.) Lag. W: A. Jasiewicz (red.), Materiały do poznania gatunków rzadkich i zagrożonych Polski. Fragmenta Floristica et Geobotanica Polonica, 36(3–4), 284–290.
Finansowanie: ze środków statutowych Zakładu Taksonomii Roślin
Potencjalny konflikt interesów: brak