

Przemysław Tomczyk*
Katedra Geobotaniki i Ekologii Roślin, Wydział Biologii i Ochrony Środowiska, Uniwersytet Łódzki
* przemyslaw.tomczyk@biol.uni.lodz.pl
Cytowanie: Tomczyk, P. (2019). Fenotypowe miary depresji wsobnej u traw – przegląd cech. Wiadomości Botaniczne, 63. https://doi.org/10.5586/wb.2019.005

Streszczenie: Wsobność (inbred, chów wsobny, krzyżowanie krewniacze) występuje wtedy, gdy w hodowli lub w naturze, krzyżują się osobniki spokrewnione ze sobą lub dochodzi do samozapylenia. Może to prowadzić do zmniejszenia różnorodności genetycznej u potomstwa poprzez zmniejszenie heterozygotyczności, a w konsekwencji do obniżonej żywotności – depresji wsobnej. Wsobność wpływa również na wzorce ekspresji genów i przykładowo obniżenie wydajności metabolizmu. Organizmy dotknięte depresją wsobną wykazują także zmiany w morfologii: osobniki są zazwyczaj mniejsze i słabsze.
Istnieje wiele sposobów na badanie depresji wsobnej i wykrywania jej. Jedną z metod jest szukanie symptomów zmniejszonego dostosowania do warunków środowiska i następnie weryfikacja genetycznego poziomu wsobności.
Wykrywanie depresji wsobnej w populacjach lub liniach hodowlanych może zapobiec dalszemu osłabianiu roślin oraz niekorzystnym zjawiskom ekologicznym lub ekonomicznym.
Celem tej pracy przeglądowej było zestawienie i porównanie najczęściej używanych i najefektywniejszych cech fenotypowych, które są przydatne do badania depresji wsobnej u traw, grupy roślin o ogromnym wpływie na ekosystemy (są ważnymi składnikami wielu biomów) i gospodarkę (jako rośliny zbożowe lub pastewne) na całym świecie. Przedstawiono przegląd 16 cech podawanych w literaturze oraz zalety i wady związane z ich wykorzystaniem, uwzględniając możliwe trudności pomiarowe. Zaprezentowano również przykłady badań, w których wykorzystano te cechy.
Phenotypic measurement of inbreeding depression in grasses – an overview of traits
Abstract: Inbreeding, in nature or cultivation, occurs when only related individuals take part in mating or in case of self-pollination. It may lead to reduced genetic diversity in the offspring by heterozygosity decrease and, in consequence, to fitness reduction – inbreeding depression. Inbreeding also influences expression patterns of genes, e.g.: inbreeding specimens can have changed (less efficient) metabolism. Organisms affected by inbreeding depression have also changed morphology: specimens are usually smaller and weaker.
There are many ways to research and detect inbreeding depression in plants. One of the methods is to find the symptoms of reduced fitness in particular plant and to check genetically the level of inbreeding in population.
Detecting of inbreeding depression in populations or breeding lines could prevent further frailty of plants, and ecological or economic defeats.
The purpose of this review was to collect and compare the most commonly used and the most effective phenotypic traits that are useful in inbreeding depression research in grasses. This group of plants significantly influences ecosystems (grasses are important compounds of many biomes) and human economy (as cereals or pastures plants) all over the world. This work presents review of 16 features from the literature, their advantages, disadvantages, and possible measurement difficulties. Examples of studies in which these features were used were also presented.
Keywords: morphology; physiology; fitness; genetic diversity; research methodology
Wsobność i depresja wsobna
Wsobność (inbred, chów wsobny, krzyżowanie krewniacze) jest procesem kojarzenia się ze sobą spokrewnionych osobników (Ryc. 1). Mamy z nim do czynienia również w przypadku samozapylenia. Może występować zarówno w naturze, w małych izolowanych populacjach, jak i hodowli roślin i zwierząt. Taki system kojarzeń prowadzi z czasem do spadku różnorodności genetycznej wśród osobników potomnych, a w konsekwencji do zmniejszonego dostosowania, czyli depresji wsobnej.
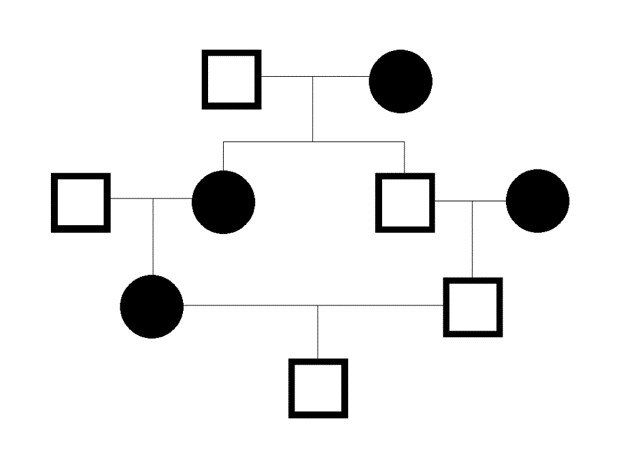
Ryc. 1. Wsobność przedstawiona na drzewie genealogicznym. Osobnik na samym dole drzewa pochodzi z chowu wsobnego, ponieważ jego rodzice byli blisko spokrewnieni. W przypadku roślin samopylnych istnieje jeszcze jeden mechanizm wsobności: może dojść do połączenia gamet z tego samego osobnika.
Istnieją dwie klasyczne hipotezy wyjaśniające przyczyny występowania depresji wsobnej: częściowej dominacji i naddominacji. Pierwsza zakłada, że chów wsobny może prowadzić do ujawniania się w homozygotach rzadkich i szkodliwych alleli recesywnych. Z kolei hipoteza naddominacji głosi wyższe dostosowanie heterozygot w porównaniu do homozygot – wzrost homozygotyczności powoduje spadek dostosowania (Kristensen i in., 2010). Spadek ten może przejawiać się w zmienionym (mniej efektywnym) metabolizmie (Kristensen i in., 2010), obniżonej odporności na stres czy zmienionej morfologii: osobniki nim dotknięte zwykle są mniejsze i słabsze (Ryc. 2) (Heschel, Paige, 1995).

Ryc. 2. Objawy wsobności mogą być bardzo wyraźne. Po prawej i lewej stronie każdego zdjęcia widzimy osobniki z 2 linii wsobnych kukurydzy. Pośrodku obu zdjęć przedstawiono osobniki niedotknięte wsobnością, o większej różnorodności genetycznej. Osobniki z linii wsobnych są m.in. znacznie niższe i mają mniejsze kolby z mniejszą liczbą nasion (źródło: seedquest.com).
Istnieją pewne wyjątki od niekorzystnych skutków wsobności. Człowiek może stosować chów wsobny do wzmacniania i ulepszania pożądanych przez siebie cech. W takim wypadku jednak krzyżowane osobniki muszą być starannie dobierane. Ponadto podczas wyprowadzania takiej ukierunkowanej linii wsobnej, co prawda część korzystnych cech może istotnie zostać utrwalona w potomstwie, jednakże będą masowo ujawniać się cechy niekorzystne, większość potomstwa w pierwszych pokoleniach może być poważnie upośledzona (Martyniuk, 2011; Siekierska, 2012). Natomiast w dzikich populacjach może zajść zjawisko oczyszczania genetycznego (ang. purging), czyli redukcja alleli niekorzystnych, oczyszczanie populacji z jej obciążenia genetycznego (Mitka, 1997).
Wsobność i depresja wsobna mogą wystąpić u wszystkich organizmów żywych rozmnażających się płciowo, także u ludzi. Przykłady tych zjawisk spotykamy np. w niektórych rodzinach królewskich, jak Habsburgowie hiszpańscy czy egipska XVIII dynastia. Oba rody upadły z powodu nagromadzenia wad genetycznych po kilku pokoleniach małżeństw między bliskimi krewnymi (Ceballos, Álvarez, 2013; Rühli, Ikram, 2014).
Depresja wsobna a fenotyp
Zjawisko depresji wsobnej zazwyczaj przekłada się na fenotyp organizmów nią dotkniętych. W roślinach efekty depresji wsobnej można zaobserwować np. w plonie nasion, zdolności kiełkowania, odporności na stres (Heschel, Paige, 1995).
Wykorzystanie fenotypowych miar depresji wsobnej jako obiektu badawczego ma rozliczne zastosowania. Na przykład za pomocą metody GWAS (ang. genome-wide association study – badanie asocjacyjne całego genomu) można powiązać określone cechy fenotypowe z ich molekularną przyczyną. W badaniu tym analizuje się zestaw wybranych cech fenotypowych z sekwencjami całych genomów badanych organizmów. Na podstawie powtarzalności współwystępowania danych wariantów SNP (ang. single-nucleotide polymorphism – polimorfizm pojedynczych nukleotydów) z daną cechą u określonych osobników można wnioskować, które geny zaangażowane są w ekspresję badanej cechy (Pearson, Manolio, 2008). Daje nam to możliwość zdobycia wiedzy, dla rozwoju których cech poziom zróżnicowania genetycznego jest szczególnie istotny, i które cechy/mechanizmy fizjologiczne będą szczególnie zagrożone w przypadku depresji wsobnej.
Badania fenotypowych miar depresji wsobnej mogą być też dobrą metodą wstępnej oceny kondycji genetycznej osobników i populacji, np. określenia które z nich mogą być dotknięte depresją wsobną, lub u których z nich może ona wystąpić w niedalekiej przyszłości. Dzięki temu możliwe jest przybliżone oszacowanie, dla których osobników warto przeprowadzić badania genetyczne ostatecznie potwierdzające poziom zróżnicowania genetycznego (Kowalczyk i in., 2018) lub które z nich będą nadawały się do danego modelu badawczego. Ponieważ nie każda placówka badawcza jest w stanie samodzielnie przeprowadzać badania genetyczne (np. brak zaplecza sprzętowego), możliwość wykonania takich doświadczeń jest pomocna.
Znaczenie traw w przyrodzie i dla człowieka
W niniejszej pracy przyglądamy się bliżej właśnie trawom z uwagi na ich wielkie znaczenie, zarówno w przyrodzie, jak i dla gospodarki. Rośliny te są ważne w naturze z kilku powodów. Są szeroko rozpowszechnione na całym świecie, nawet na Antarktydzie. Spośród dwóch występujących tam rodzimych gatunków roślin naczyniowych, jeden jest trawą (śmiałek antarktyczny Deschampsia antarctica E. Desv). Ponadto występuje tam rozprzestrzeniona przez człowieka wiechlina roczna Poa annua L. (Chwedorzewska, 2008; Parnikoza i in., 2007). W cieplejszych strefach klimatycznych trawy mogą tworzyć ekosystemy, w których są gatunkami dominującymi: stepy, sawanny, prerie, pampasy, campos czy puszta. Zajmują one blisko 1/3 powierzchni lądów, a pod względem produkcji biomasy ustępują tylko lasom. Trawy są również rodziną bardzo bogatą w rodzaje i gatunki, ustępują pod tym względem tylko Asteraceae, Fabaceae i Orchidaceae. Wreszcie, są bardzo zróżnicowane morfologicznie, od drobnych kilkucentymetrowych roślin zielnych po kilkunastometrowe bambusy z drewniejącymi łodygami (Mitka, 1997).
Trawy odgrywają także doniosłą rolę w rolnictwie. Wszystkie zboża należą do tej grupy roślin, a ich znaczenie w produkcji żywności i farmaceutyce jest nieocenione (Frey, 2010). Trawy są ważne również w chowie zwierząt gospodarskich, stanowiąc dla nich paszę. Tak więc badania nad trawami mogą silnie przekładać się na korzyści w rolnictwie i ochronie środowiska naturalnego.
Biorąc pod uwagę znaczenie traw i opisane wcześniej zjawisko depresji wsobnej, za warte uwagi uznano przyjrzenie się możliwościom wstępnej fenotypowej oceny kondycji genetycznej traw i odpowiedź na pytanie: które cechy fenotypowe są użyteczne do badania depresji wsobnej u tych roślin? Przegląd tych cech przedstawiono w dalszej części pracy.
Zasady planowania eksperymentu
Planując prowadzenie badań nad depresją wsobną i jej fenotypowymi miarami należy pamiętać o kilku zasadach. Aby wykluczyć działanie plastyczności fenotypowej i wpływu środowiska na cechy fenotypowe, badane rośliny powinny rosnąć w takich samych warunkach (wspólna uprawa ogrodowa, ang. common garden experiment, lub uprawa w szklarni czy fitotronie). Najlepszym rozwiązaniem byłoby wysianie nasion w tym samym czasie, w celu dalszej wspólnej uprawy. Kiedy takie rozwiązanie byłoby trudne lub niemożliwe w realizacji, do uprawy można przeznaczyć jednakowej wielkości sadzonki roślin (np. w przypadku traw wieloletnich – fragmenty kęp). Należy w takiej sytuacji zaczekać z pomiarami do kolejnego sezonu wegetacyjnego lub nawet dwa sezony (Gibson, Nelson, 2017).
Odnośnie samych pomiarów cech fenotypowych, powinno je się prowadzić w tym samym czasie, o ile to możliwe tego samego dnia. Aby wykluczyć sezonową zmienność roślin wieloletnich, pomiary powinny być powtórzone w minimum dwóch sezonach badawczych. W przypadku roślin jednorocznych takie eksperymenty prowadzone są zwykle przez jeden sezon. Dlatego istotne jest, aby każdy element modelu w doświadczeniu (np. populacji, ekotypu czy odmiany) był odpowiednio licznie reprezentowany dla wystarczającej próby statystycznej (Yadav i in., 2018).
Zasady przeprowadzania eksperymentu są takie same dla roślin uprawnych i dla form dzikich. W przypadku form dzikich należy pamiętać o ewentualnych zezwoleniach, jeżeli jest to gatunek chroniony oraz o środkach ostrożności jeśli to gatunek inwazyjny. Z kolei w przypadku form uprawnych należy uwzględnić odpowiednie procedury, jeżeli jest to gatunek inwazyjny dla danego terenu, lub forma modyfikowana genetycznie (de Villemereuil i in., 2016; Gallagher i in., 1988; Thompson i in., 1994).
Przegląd cech
Na podstawie przeglądu literatury zebrano 16 cech, które można zastosować do badań depresji wsobnej traw (Alam i in., 2004; Keller, Waller, 2002; Oakley, Winn, 2008; Tyler i in., 2014). Każdej z nich przyporządkowano zalety i wady związane z zastosowaniem wybranych cech do pomiarów, dla każdej podano także przykładowe źródła literaturowe zawierające wykorzystanie danej cechy w badaniach związanych z wsobnością. Dane przedstawiono w tabeli 1.
Przy określaniu zalet danej cechy brano pod uwagę:
- łatwość pomiaru – możliwość szybkiego i sprawnego zebrania wyników, bez konieczności posiadania specjalistycznego sprzętu;
- możliwość uzyskania bardzo dużej próby statystycznej z osobnika (większa rozdzielczość wyników) – co pozwala na uzyskanie bardziej wiarygodnego wyniku uśrednionego;
- dane na temat dynamiki rozwoju roślin – dające wgląd w tempo rozwoju rośliny: jak szybko się rozwija, jaka jest ilość czasu potrzebna na realizację danej fazy rozwoju?;
- dostarczanie bezpośrednich informacji o fizjologii (żywotności) osobników – cechy pozwalające ocenić reakcję rośliny na bodźce środowiska, a nie tylko na efekt końcowy (np. różnice w morfologii). Dzięki temu uzyskujemy pełniejsze tło i możliwe są wnioski dotyczące gorszego rozwoju osobników i ewentualnych sposobów poprawy tego niekorzystnego stanu.
Z kolei w przypadku wad uwzględniano:
- pracochłonność – konieczność wykonania bardzo wielu pomiarów. Uzyskane dane mają zwykle dużą wartość, z uwagi na dużą próbę statystyczną, jednak ich pozyskanie może być uciążliwe;
- czasochłonność – konieczność monitorowania upraw przez dłuższy czas (aby wychwycić daną fazę rozwoju rośliny);
- fakt, iż osobniki potomne (nasiona) są genetycznymi hybrydami rośliny matecznej i tej, z której pochodził pyłek. Aby uzyskane wyniki nie były zniekształcone, podczas kwitnienia należy zadbać, aby nie dochodziło do krzyżowego zapylenia między osobnikami z różnych linii/populacji;
- sezonowość owocowania: zbiór nasion musi odbyć się w odpowiednim czasie, należy monitorować stan ich dojrzałości (ryzyko osypania się części nasion i zniekształcenia wyników);
- konieczność stosowania specjalistycznego sprzętu lub zaplanowania złożonego eksperymentu – w pierwszym wypadku pojawia się problem kosztów, na które nie każdy zespół badawczy może sobie pozwolić, w drugim przypadku doświadczenie będzie długotrwałe.
W przypadku każdego gatunku trawy zakres normy badanej cechy będzie inny. Dlatego, aby stwierdzić czy dany osobnik jest dotknięty depresją wsobną, potrzebne są dane literaturowe na temat zakresu wielkości danej cechy lub porównanie z osobnikami tego samego gatunku (bądź odmiany czy ekotypu w przypadku dużego zróżnicowania w obrębie gatunku) o dobrej kondycji genetycznej. Istotna statystycznie różnica pomiędzy wartościami cechy, na niekorzyść badanego osobnika, może świadczyć o symptomach depresji wsobnej.
Wybór konkretnych cech do zastosowania w badaniach będzie uzależniony od specyfiki prowadzonych doświadczeń. Z tego powodu nie podawano cech „najlepszych”, ponieważ dla każdego zespołu badawczego argumenty przemawiające na korzyść cechy mogą być inne (np. finanse, czas, zaplecze badawcze, liczebność personelu). Ograniczono się do podania wad i zalet każdej cechy. Planując eksperyment zawsze warto wybrać co najmniej kilka cech do zbadania, aby uzyskać pełniejszy obraz problemu. O ile pozwalają na to warunki, warto zbadać nie tylko cechy morfologiczne, ale uwzględnić również cechy związane z fizjologią oraz dynamiką rozwoju roślin. Dzięki temu uzyskuje się wgląd w większą liczbę aspektów życia i funkcjonowania rośliny. Ponadto umożliwia to porównywanie ze sobą różnych badań (Sandner, 2016).
Tab. 1. Cechy fenotypowe użyteczne do badania depresji wsobnej u traw.
| Cecha | Zalety | Wady i trudności | Przykłady źródeł wymieniających daną cechę |
|---|---|---|---|
| Liczba źdźbeł | Łatwość pomiaru | – | Keller, Waller, 2002, s.234 |
| Długość źdźbeł | Łatwość pomiaru | – | Alam i in., 2004, s. 409
Erickson i in., 2004, s. 1780 Knapp, Rice, 2008 Tyler i in., 2014, s. 1 Yadav i in., 2018, s. 615 |
| Liczba kłosków na źdźble | Możliwość uzyskania bardzo dużej próby statystycznej z osobnika (większa rozdzielczość wyników) | Czaso- i pracochłonne | Alam i in., 2004, s. 409
Torkian i in. 2018, s. 568 Yadav i in., 2018, s. 615 |
| Procent kiełkujących nasion | Możliwość uzyskania bardzo dużej próby statystycznej z osobnika rodzicielskiego (większa rozdzielczość wyników)
Dostarcza bezpośrednich informacji o fizjologii (żywotności) osobników |
Czaso- i pracochłonne (zwłaszcza w przypadku nasion wymagających stratyfikacji)
W przypadku gatunków obcopylnych osobniki potomne (nasiona) są genetycznymi hybrydami rośliny matecznej i tej, z której pochodził pyłek. Aby uzyskane wyniki nie były zniekształcone, podczas kwitnienia, należy zadbać aby nie dochodziło do krzyżowego zapylenia między osobnikami z różnych linii/populacji |
Erickson i in., 2004, s. 1780
Keller, Waller, 2002, s.236 Oakley, Winn, 2008, s. 527 |
| Masa ziarniaków (np. masa 100 lub 1000 ziarniaków) | Łatwość pomiaru | W przypadku gatunków obcopylnych osobniki potomne (nasiona) są genetycznymi hybrydami rośliny matecznej i tej, z której pochodził pyłek. Aby uzyskane wyniki nie były zniekształcone, podczas kwitnienia, należy zadbać aby nie dochodziło do krzyżowego zapylenia między osobnikami z różnych linii/populacji | Alam i in., 2004, s. 409
Erickson i in., 2004, s. 1780 Knapp, Rice, 2008 Oakley, Winn, 2008, s. 527 Torkian i in. 2018, s. 568 Tyler i in., 2014, s. 1 Yadav i in., 2018, s. 615 |
| Długość ziarniaków | Możliwość uzyskania bardzo dużej próby statystycznej z osobnika (większa rozdzielczość wyników) | W przypadku gatunków obcopylnych osobniki potomne (nasiona) są genetycznymi hybrydami rośliny matecznej i tej, z której pochodził pyłek. Aby uzyskane wyniki nie były zniekształcone, podczas kwitnienia, należy zadbać aby nie dochodziło do krzyżowego zapylenia między osobnikami z różnych linii/populacji | Tyler i in., 2014, s. 1 |
| Szerokość ziarniaków | Możliwość uzyskania bardzo dużej próby statystycznej z osobnika (większa rozdzielczość wyników) | W przypadku gatunków obcopylnych osobniki potomne (nasiona) są genetycznymi hybrydami rośliny matecznej i tej, z której pochodził pyłek. Aby uzyskane wyniki nie były zniekształcone, podczas kwitnienia, należy zadbać aby nie dochodziło do krzyżowego zapylenia między osobnikami z różnych linii/populacji | Keller, Waller, 2002, s. 236
Tyler i in., 2014, s. 1 |
| Liczba ziarniaków na źdźbło (lub z określonej liczby źdźbeł, np. 10) | Możliwość uzyskania bardzo dużej próby statystycznej z osobnika (większa rozdzielczość wyników) | Czaso- i pracochłonne.
Zbiór nasion musi odbyć się w odpowiednim czasie (ryzyko osypania się części nasion i zniekształcenia wyników) |
Alam i in., 2004, s. 409
Erickson i in., 2004, s. 1780 Keller, Waller, 2002, s.236 Knapp, Rice, 2008 Oakley, Winn, 2008, s. 527 |
| Odporność na stres (np. pomiar poziomu hormonów stresu podczas suszy lub śmiertelność i różnice we wzroście po epizodzie zgryzania przez roślinożerców) | Dostarcza bezpośrednich informacji o fizjologii (żywotności) osobników i ich zdolnościach adaptacyjnych. | Czaso- i pracochłonne.
Wymaga specjalistycznego sprzętu lub zaplanowania złożonego eksperymentu |
Keller, Waller, 2002, s.238
Kozub i in. 2017, s.262 |
| Wysokość rośliny | Łatwość pomiaru | – | Alam i in., 2004, s. 409
Tyler i in., 2014, s. 1 |
| Długość liścia flagowego | Łatwość pomiaru | – | Alam i in., 2004, s. 409
Erickson i in., 2004, s. 1780 Knapp, Rice, 2008 Torkian i in. 2018, s. 568 |
| Liczba dni do rozpoczęcia fazy liścia flagowego | Dane na temat dynamiki rozwoju roślin | Czasochłonne | Alam i in., 2004, s. 409
|
| Liczba dni do rozpoczęcia fazy wiechy (lub innego typu kwiatostanu – zależnie od gatunku) | Dane na temat dynamiki rozwoju roślin | Czasochłonne | Alam i in., 2004, s. 409
|
| Liczba dni do 100 % kwitnienia | Dane na temat dynamiki rozwoju roślin | Czasochłonne | Alam i in., 2004, s. 409
Yadav i in., 2018, s. 615 |
| Liczba dni do dojrzałości ziarniaków | Dane na temat dynamiki rozwoju roślin. | Czasochłonne. | Alam i in., 2004, s. 409
|
| Liczba plennych kłosków/źdźbło | Łatwość pomiaru | Czaso- i pracochłonne | Alam i in., 2004, s. 409
|
Podsumowanie – przykłady zastosowań fenotypowych miar depresji wsobnej
Badanie cech fenotypowych nadal pozostaje dobrym sposobem oceny dostosowania osobników i populacji, pozwalając wysnuć pewne wnioski o ich kondycji genetycznej: cechy fenotypowe w większości warunkowane są wieloma genami, a przez to każda cecha niesie informacje o wielu loci w całym genomie (Richards i in., 2016). Ponadto udowodniono, że populacje bardziej jednorodne fenotypowo (a więc i genotypowo) są znacznie bardziej narażone na wymarcie (Richards i in., 2016). Ponieważ cechy fenotypowe są bezpośrednio związane z dostosowaniem do danego środowiska, bywają wykorzystywane w ochronie przyrody: do wyboru optymalnych osobników w restytucji gatunków, programach konserwatorskich czy rekultywacji (Richards i in., 2016).
Interesujące badania na ten temat przeprowadzono np. dla Nasella pulchra (Hitchc.) Barkworth (Knapp, Rice, 2008). W pracy zbadano zarówno cechy molekularne (izozymy) jak i fenotypowe roślin z kilku różnych populacji. Wśród wziętych pod uwagę cech morfologicznych były m.in.: wysokość źdźbła, długość liścia, liczba ziarniaków ze źdźbła i masa ziarniaków. Obie klasy cech (molekularne i fenotypowe) korelowały ze sobą, ale pojawiały się także różnice – jedynie cechy fenotypowe były związane z czynnikami klimatycznymi. Autorzy wskazali na zalety korzystania z cech morfologicznych. Dzięki nim, korzystając z łatwo dostępnych danych klimatycznych, możemy przewidzieć, które osobniki będą lepiej przystosowane do wzrostu na danym terenie. Na podstawie fenotypu jesteśmy bowiem w stanie stwierdzić, do jakiego ekotypu należą osobniki i które tereny będą dla nich odpowiedniejsze. W przypadku osobników dotkniętych depresją wsobną to szczególnie istotne.
Podobne doświadczenia wykonano dla Elymus glaucus Buckley (Erickson i in., 2004). Głównym celem badania było znalezienie wzorców geograficznych adaptacyjnego zróżnicowania tej trawy. Takie informacje można wykorzystać do planowania dystrybucji nasion i zachowania adaptacyjnych wzorców zmienności genetycznej w trwających pracach konserwatorskich. Większość cech morfologicznych jakich użyto w tym projekcie pokrywa się z poprzednio opisanym przypadkiem, były to m.in.: długość liści, wysokość źdźbeł, liczba nasion ze źdźbła.
Innym ciekawym przykładem zastosowania fenotypowych miar depresji wsobnej są doświadczenia na życicy trwałej Lollium perenne L. (Torkian i in., 2019). Badano tam wpływ obecności grzyba endofitycznego na rośliny o niskiej różnorodności genetycznej. Jako fenotypowych wskaźników wsobności użyto m.in. długość liści, liczbę kłosków na źdźble i masę nasion. W badaniu wykazano, że endofity mogą wydatnie zmniejszać fenotypowe efekty depresji wsobnej.
Badania nad związkiem genotypu i fenotypu prowadzi się od dekad. Dla ludzi szczególnym obiektem zainteresowania są te cechy, które będziemy mogli wykorzystać do naszego bezpośredniego użytku, jak masa ziarniaków czy odporność na niekorzystne, stresowe warunki. Doświadczenia nad Trichloris crinita (Lag.) Parodi. prowadzono z myślą o wykorzystaniu jej w Ameryce Północnej podczas rekultywacji zdegradowanych terenów, czy jako cenne źródło paszy dla zwierząt (Kozub i in., 2017). W badaniach, poza cechami morfologicznymi, analizowano również odporność na stres, jako ważną zaletę w rekultywacji zdewastowanych terenów. Badania nad fenotypem tych roślin i ich powiązanie z kondycją genetyczną populacji pomogą zaplanować efektywniejsze programy hodowlane i ulepszyć cechy gatunku dla naszych korzyści.
Również w przypadku jęczmienia zwyczajnego Hordeum vulgare L. badano różnorodność genetyczną, z uwzględnieniem poziomu depresji wsobnej oraz fenotypu roślin (m.in.: liczbę kłosków, masę ziarniaków, wysokość roślin i liczbę dni potrzebną do zakwitnięcia) (Yadav i in., 2018). Doświadczenia wykazały, że cecha najbardziej użyteczna dla ludzi, plon ziarniaków, jest skorelowana z pozostałymi cechami morfologicznymi użytymi w analizie, zatem w programach hodowlanych odmian, je również można wykorzystywać podczas selekcji.
Z kolei na Zoysia matrella (L.) Merr. badano wpływ samozapylenia (i pojawienia się depresji wsobnej) na cechy morfologiczne. Autorzy stwierdzili, że samozapylenie można zastosować do selekcji pożądanych cech z wysoce heterozygotycznych gatunków (jak Z. matrella). Ponadto wyprowadzone przez autorów linie hodowlane mogą zostać wykorzystane w dalszych badaniach ilościowej analizy cech morfologicznych (Kunwanlee i in., 2017).
Praktyczne zastosowanie znalazły również badania nad Paspalum dilatatum subsp. flavescens Roseng., Arrill. & Izag. Analiza danych genetyczynych i fenotypowych wykazała, że populacje położone na skraju zasięgu były mniej zróżnicowane genetycznie oraz różniły się morfologicznie od innych – populacje te będą cennym materiałem w hodowli odmian tej trawy (SandroGutiérrez, Speranza, 2019).
Jak widać po przytoczonych przykładach, mimo, że badanie cech fenotypowych depresji wsobnej może czasem wydawać się metodą tradycyjną, nie tak zaawansowaną jak np. metody molekularne, to pozostaje ono ciągle cennym narzędziem, które w połączeniu z innymi metodami umożliwia dokonywanie wartościowych i użytecznych odkryć.
Literatura
Alam, M. F., Khan, M. R., Nuruzzaman, M., Parvez, S., Swaraz, A. M., Alam, I., & Ahsan, N. (2004). Genetic basis of heterosis and inbreeding depression in rice (Oryza Sativa L.). Journal of Zhejiang University-SCIENCE A, 5(4), 406–411. https://doi.org/10.1631/jzus.2004.0406
Ceballos, F., & Álvarez, C. G. (2013). Royal dynasties as human inbreeding laboratories: the Habsburgs. Heredity, 111(2), 114–121. https://doi.org/10.1038/hdy.2013.25
Chwedorzewska, K. J. (2008). Poa annua L. in Antarctic: searching for the source of introduction. Polar Biology, 31(3), 263–268. https://doi.org/10.1007/s00300-007-0353-4
Erickson, V. J., Mandel, N. L., & Sorensen, F. C. (2004). Landscape patterns of phenotypic variation and population structuring in a selfing grass, Elymus glaucus (blue wildrye). Canadian Journal of Botany, 82(12), 1776–1789 . https://doi.org/10.1139/b04-141
Frey, L. (2010). Grasses in Poland: invincible, but threatened. Biodiversity Research and Conservation, 19(1), 93–102. https://doi.org/10.2478/v10119-010-0025-z
Gallagher, J. L., Somers, G. F., Grant, D. M., & Seliskar, D. M. (1988). Persistent differences in two forms of Spartina alterniflora: a common garden experiment. Ecology, 69(4), 1005–1008. https://doi.org/10.2307/1941255
Gibson, A., & Nelson C. R. (2017). Comparing provisional seed transfer zone strategies for a commonly seeded grass, Pseudoroegneria spicata. Natural Areas Journal, 37, 188–199. https://doi.org/10.3375/043.037.0208
Heschel, M. S., & Paige, K. N. (1995). Inbreeding depression, environmental stress, and population size variation in scarlet gilia (Ipomopsis aggregata). Conservation Biology, 9(1), 126–133. https://doi.org/10.1046/j.1523-1739.1995.09010126.x
Keller, L. F., & Waller, D. M. (2002). Inbreeding effects in wild populations. Trends in Ecology & Evolution, 17(5), 230–241. https://doi.org/10.1016/S0169-5347(02)02489-8
Knapp, E. E., & Rice, K. J. (2008). Comparison of isozymes and quantitative traits for evaluating patterns of genetic variation in purple needlegrass (Nassella pulchra). Conservation Biology, 12, 1031–1041. https://doi.org/10.1046/j.1523-1739.1998.97123.x
Kowalczyk, M., Szabelak, A., Dylewska, M., & Jakubczak, A. (2018). Markery molekularne wykorzystywane w selekcji zwierząt hodowlanych. Zeszyty Problemowe Postępów Nauk Rolniczych, 2018(592), 37–49. https://doi.org/10.22630/ZPPNR.2018.592.4
Kozub, P. C., Cavagnaro, J. B., & Cavagnaro, P. F. (2017). Exploiting genetic and physiological variation of the native forage grass Trichloris crinita for revegetation in arid and semi‐arid regions: an integrative review. Grass and Forage Science, 73(2), 257–271. https://doi.org/10.1111/gfs.12337
Kristensen, T. N., Pedersen, K. S., Vermeulen, C. J., & Loeschcke, V. (2010). Research on inbreeding in the “omic” era. Trends in Ecology & Evolution, 25(1), 44–52. https://doi.org/10.1016/j.tree.2009.06.014
Kunwanlee, P., Tanaka, H., Hashiguchi, M., Gondo, T., Muguerza, M., Inoue, T., & Akashi, R. (2017). The highly heterozygous homoploid turfgrass Zoysia matrella displays desirable traits in the S1 progeny. Crop Science, 57, 3310–3318. https://doi.org/10.2135/cropsci2016.09.0811
Martyniuk, E. (2011). Problemy hodowlane populacji o małej liczebności na przykładzie owcy rasy olkuskiej. Wiadomości Zootechniczne, 49(1), 21–31.
Mitka, J. (1997). Małe, izolowane populacje na skraju zasięgu geograficznego – niektóre procesy ekologiczne i genetyczne. Wiadomości Botaniczne, 41(2), 13–34.
Oakley, C. G., & Winn, A. A. (2008). Population‐level and family‐level inbreeding depression in a cleistogamous perennial. International Journal of Plant Sciences, 169(4), 523–530. https://doi.org/10.1086/528752
Parnikoza, I. Y., Maidanuk, D. N., & Kozeretska, I. A. (2007). Are Deschampsia antarctica Desv. and Colobanthus quitensis (Kunth) Bartl. migratory relicts? Cytology and Genetics, 41(4), 226–29. https://doi.org/10.3103/S0095452707040068
Pearson, T. A., & Manolio, T. A. (2008). How to Interpret a Genome-Wide Association Study. JAMA, 299 (11), 1335–44. doi: 10.1001/jama.299.11.1335.
Richards, C.M., Falk, D. A., & Montalvo, A. M. (2016). Population and ecological genetics in restoration ecology. W: M. A. Palmer, J. B. Zedler, & D. A. Falk (red.), Foundations of Restoration Ecology (pp. 123–152). Washington D.C.: Island Press. https://doi.org/10.5822/978-1-61091-698-1_5
Rühli, F. J., Ikram, S. (2014). Purported medical diagnoses of Pharaoh Tutankhamun, c. 1325 BC-. HOMO, 65(1), 51–63. https://doi.org/10.1016/j.jchb.2013.08.006
Sandner, T. M. (2016). The effects of inbreeding and stress on plant performance [Rozprawa doktorska, Philipps-Universität Marburg]. Marburger Publikationsserver. https://archiv.ub.uni-marburg.de/diss/z2016/0080/pdf/dts.pdf
Sandro, P., Gutiérrez, L., & Speranza, P. (2019). Distribution of genetic and phenotypic diversity in the autogamous perennial Paspalum dilatatum subsp. flavescens Roseng., Arrill. & Izag. (Poaceae). Genetic Resources and Crop Evolution, 66, 1205. https://doi.org/10.1007/s10722-019-00791-9
Siekierska, A. (2012). Inbred w hodowli bydła musi być kontrolowany. Przegląd hodowlany, 2012(7–9), 23–24.
Thompson, J. D., Higgins, D. G., & Gibson, T. J. (1994). CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Research, 22(22), 4673–4680. https://doi.org/10.1093/nar/22.22.4673
Torkian, M., Sabzalian, M. R., & Ehtemam, M. H. (2019). A simultaneous effect of selfing and Epichloë endophyte on forage, seed yield and turf characteristics of perennial ryegrass (Lolium perenne L.). Grass and Forage Science, 74, 559–570. https://doi.org/10.1111/gfs.12437
Tyler, L., Fangel, J. U., Fagerström, A. D., Steinwand, M. A., Raab, T. K., Willats, W. G. T., & Vogel, J. P. (2014). Selection and phenotypic characterization of a core collection of Brachypodium distachyon inbred lines. BMC Plant Biology, 14(1). https://doi.org/10.1186/1471-2229-14-25
Villemereuil, P. de, Gaggiotti, O. E., Mouterde, M., & Till-Bottraud, I. (2016). Common garden experiments in the genomic era: new perspectives and opportunities. Heredity, 116(3), 249–254. https://doi.org/10.1038/hdy.2015.93
Yadav, H. C., Singh, S. K., Gupta, P. K., Yadav, P. C., & Chaurasiya, J. P. (2018). Studies on path coefficient analysis and genetic divergence in feed barley (Hordeum vulgare L.). Journal of Pharmacognosy and Phytochemistry, 7(2), 613–616.
Finansowanie: Praca powstała w ramach realizacji grantu finansowanego przez Narodowe Centrum Nauki, grant PRELUDIUM 12, nr: 2016/23/N/NZ8/02057, „Poszukiwanie epigenetycznych różnic we wzorcach reakcji na stres środowiskowy w populacjach izolowanych i otwartych Festuca tatrae i Festuca amethystina”.
Potencjalny konflikt interesów: brak